
These data most likely reflect the difference between AAV1 and AAV6 interactions with sialic acid during the initial binding and cell entry steps. Industry is dominated and accounted for the major income share In 2022. The major market players are taking enormous steps to rise their market position through developing countries, including increasing research and development investments and constant mergers and acquisitions. Market Reports World is the Credible Source for Gaining the Market Reports that will Provide you with the Lead Your Business Needs. With this information, stakeholders will be more capable of developing new strategies, which focus on market opportunities that will benefit them, making their business endeavors profitable in the process. What are the key market trends impacting the growth of the Sialic Acid market? AAV6 binding appears to be less affected by neuraminidase compared with AAV1 binding, suggesting that AAV6 may also bind to moieties other than sialic acid on the cell surface. Consistent with the results shown in Fig. Fig.22 and and3,3, transduction by AAV6 appears to be more dependent on sialic acid than AAV1. Similar to the neuraminidase treatment experiment (Fig. (Fig.2D),2D), a larger amount of AAV6 bound to the Lec-2 cells compared to AAV1, consistent with its possible utilization of additional carbohydrates in binding to these cells.
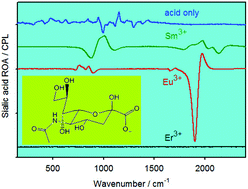
 In contrast, neuraminidase treatment markedly decreased gene transduction by either AAV1 or AAV6 vectors on all the cells tested (Fig. 2A to C). The sialic acid that facilitates AAV1 and AAV6 transduction localizes on glycoproteins rather than on glycolipids. Previous studies have shown that both AAV4 and AAV5 use α2,3 sialic acid for efficient binding and transduction (19). We asked if AAV1 and AAV6 have a preference for a sialic acid linkage(s) for efficient transduction. As shown in Fig. Fig.6,6, for both cells lines, proteinase K treatment inhibited more than 80% of transduction by AAV1, AAV6, and AAV5, suggesting that the receptors of these viruses are glycoproteins. Treatment of Lec-2 cells with either sialyltransferase or CMP-sialic acid alone did not result in successful resialylation and any increased transduction by the viruses (data not shown). AAV2 binding to Pro-5 cells was not significantly reduced after neuraminidase treatment. MAA blocked AAV1 and AAV6 transduction on Pro-5 and Cos-7 cells, which display α2,3 sialic acid on their surfaces, but not on HepG2 cells, which display α2,6 sialic acid. There was a 12- or 98-fold decrease in AAV6 transduction following neuraminidase treatment on HepG2 and Pro5 cells, respectively, in contrast to a 5- or 37-fold decrease in AAV1 transduction on these two cell lines.
In contrast, neuraminidase treatment markedly decreased gene transduction by either AAV1 or AAV6 vectors on all the cells tested (Fig. 2A to C). The sialic acid that facilitates AAV1 and AAV6 transduction localizes on glycoproteins rather than on glycolipids. Previous studies have shown that both AAV4 and AAV5 use α2,3 sialic acid for efficient binding and transduction (19). We asked if AAV1 and AAV6 have a preference for a sialic acid linkage(s) for efficient transduction. As shown in Fig. Fig.6,6, for both cells lines, proteinase K treatment inhibited more than 80% of transduction by AAV1, AAV6, and AAV5, suggesting that the receptors of these viruses are glycoproteins. Treatment of Lec-2 cells with either sialyltransferase or CMP-sialic acid alone did not result in successful resialylation and any increased transduction by the viruses (data not shown). AAV2 binding to Pro-5 cells was not significantly reduced after neuraminidase treatment. MAA blocked AAV1 and AAV6 transduction on Pro-5 and Cos-7 cells, which display α2,3 sialic acid on their surfaces, but not on HepG2 cells, which display α2,6 sialic acid. There was a 12- or 98-fold decrease in AAV6 transduction following neuraminidase treatment on HepG2 and Pro5 cells, respectively, in contrast to a 5- or 37-fold decrease in AAV1 transduction on these two cell lines.
For cells, saccharides, nucleic acids, and polypeptides of the invention, the term “isolated” refers to material that is substantially or essentially free from components which normally accompany the material as found in its native state. In contrast, AAV4, which uses O-linked sialic acid for transduction, transduced Lec-1 cells fourfold more efficiently than Pro-5 cells, suggesting that removal of the N-linked glycan facilitates AAV4 interaction with O-linked glycan. Resialylation experiments confirmed that AAV1 and AAV6 use α2,3 or α2,6 N-linked sialic acid for efficient transduction. When you have any questions regarding where and tips on how to use sialic acid powder factory, it is possible to email us with the webpage. In contrast, resialylation with α2,3(O)-sialyltransferase did not affect AAV1 and AAV6 transduction, while resialylation with α2,3(N)- or α2,6(N)-sialyltransferase resulted in substantial increases in AAV1 and AAV6 transduction (Fig. (Fig.8A).8A). Therefore, Pro-5 cells and HepG2 cells display α2,3 sialic acid or α2,6 sialic acid on their surfaces, respectively, while Cos-7 cells display α2,3 sialic acid and a relatively small amount of α2,6 sialic acid. HepG2 (A) and Pro-5 (B) cells were transduced with a constant amount of AAV1-luc or AAV6-luc vectors in the presence of a 200-fold-excess amount of competing AAV1, AAV6, or AAV2 encapsidated λ phage DNA-containing vector at 37°C for 1 h. Lectin competition on HepG2 (A), Pro-5 (B), and Cos-7 (C) cells.
As a control, AAV2 transduction was not affected by lectin competition on all cell lines (data not shown). As a negative control, transduction by AAV2 did not obviously change after resialylation with each sialyltransferase (Fig. (Fig.8C).8C). Resialylation by α2,3(N)-sialyltransferase and α2,6(N)-sialyltransferase increased AAV6 transduction by 11-fold and 6-fold, respectively, but only 2.5-fold for AAV1. These results again support the idea that sialic acid facilitates AAV1 and AAV6 transduction, in particular both α2,3 and α2,6 sialic acids. We stained cell lines with lectins that have been used to recognize the following three different epitopes: (i) WGA recognizes all sialic acids, (ii) MAA recognizes α2,3 sialic acid, and (iii) SNA recognizes α2,6 sialic acid. Cultures were stained with FITC-labeled lectins that bind to three different carbohydrates as follows: WGA binds sialic acid in any linkage, MAA binds 2,3-linked sialic acid, and SNA binds 2,6-linked sialic acid. WGA, which binds to all linkage forms of sialic acid, blocked both AAV1 and AAV6 transduction on all the cells tested. Neuraminadase treatment of cells reduces AAV1 and AAV6 binding and transduction. E. coli , by calcium chloride transformation, and into eukaryotic cells by calcium phosphate treatment or electroporation. In contrast, 89% and 68% inhibition of AAV1 and AAV6 binding, respectively, was observed after neuraminidase treatment.

